

Introduction
Materials and Methods
Pollen Morphologic Characteristics
Pollen Germination Rate
Pollen Tube Elongation
Pollen Fertilization Rate and Fruit Weight
Statistical Analysis
Results and Discussion
Pollen Morphological Characteristics
Pollen Germination and Pollen Tube Elongation
Fruit-Set Rate and Fruit Weight of Pollen for Pollination
Introduction
Pear, belonging to the Rosaseae and Pomoideae families and the genus Pyrus, is common in temperate regions. Pear was cultivated on 22,567 ha in South Korea in 2007, but this area has decreased each year to 10,837 ha in 2017, a decrease of 52%. Production amounts were about 265,757 tons (Seo et al., 2011). The composition of pear tree cultivars has also changed. The cultivation area ratio of ‘Niitaka’ (P. pyrifolia cv. Niitaka) was 79.9% in 2007 and rose to 86.8% in 2017, revealing the dominance of that single variety. By contrast, the cultivation area of ‘Wonwhang’ fell slightly from 5.4 to 4.7% during the same period and that of other varieties dropped greatly from 19.6 to 8.5% over the same period (KREI, 2018).
Pears go through pollination and fertilization stages for fruit set. Pollen from the same pear variety cannot be used for fertilization, indicating self-incompatibility and highlighting the need for cross-fertilization. The major cultivar of South Korea, ‘Niitaka’, has no pollen (Seo et al., 2011). Therefore, for fruit set, at least 20% of the major cultivar must be pollinizers. With a sufficient amount of pollinizers and flower-visiting insects, natural pollination can be achieved. However, recent environmental factors, such as weather conditions and agricultural chemical spraying, have reduced the number of flower-visiting insects. Chilling injury in the flowering season, among other factors, has led to an increased use of artificial pollination (Peak and Cheong, 1977; Kang et al., 2010). In South Korea, 91.2% of the domestic pear cultivation areas and 74.3 % of the cultivating farms implement artificial pollination.
Important factors for a successful artificial pollination are known to be pollen status, artificial pollination time, weather conditions, etc. Suitable pollinizers must have the same flowering time as that of the main varieties, as well as high interspecific compatibility, pollen activity, germinative power, and pollen production (Griggs and Scharader, 1942; Crassweller et al., 1980; Cothard, 1994; Kang et al., 2010; Bieniasz et al., 2019).
The pear ovary is separated into five locules, and each locule has two ovules. If all of the ovules are fertilized, 10 seeds are generated (Bae and Kim, 2002). The impact of different pollinizer varieties on fruit quality varies according to the seed generation ability and metaxenia directly influenced by pollen (Hibbard, 1933; Tufts and Hanse, 1933). Moreover, as fruit size is in direct proportion to the number of seeds, the fruit part of ovary with poor seed formation does not develop and grows into an abnormal fruit (Shin et al., 1988; Kim et al., 2018; Kang, 2010).
The pollen presently utilized for pear artificial pollination in South Korea consists of diverse varieties, including those collected by farms (‘Chuwhangbae’ and ‘Wonwhang’) and those imported from China (‘Xuehuali’ and ‘Yali’). South Korea relies on import for about 90% of its pollen. Major pear pollen exporters are China and South East Asia. About 2,000 kg of pear pollen is imported every year (Lee et al., 2016; Lee et al., 2017). Pollen for pollination must be compatible with major cultivars, so its varieties should be accurately identified. It is difficult to differentiate varieties just by looking at pollen, although systematic botany has used pollen taxonomy (Lee, 1986) based on the pollen’s morphological characteristics and surface pattern (Thakur and Thakur, 1970). Shim et al. (1988a) compared the patterns on pear pollen surfaces and found that Japanese pear had almost the same width and linear of striate bumps, while Chinese pear had narrower widths than Japanese pear. Also, the distance between striate bumps was wider in Chinese pear than in Japanese pear.
Diverse domestic and imported pollen varieties are utilized in China for pear pollination. However, yearly fertilization rates are different possibly due to environmental factors and pollen defects.
Against this backdrop, this study investigated domestic pollen and imported pollen from China for pear pollination to determine their morphological characteristics and germination and fertilization abilities. This study provides the basic materials for identifying pollen-related causes of defects in pear fertilization.
Materials and Methods
Pollen Morphologic Characteristics
Five pollen varieties frequently utilized for pear pollination in South Korea were studied in this experiment: ‘Xuehuali’ and ‘Yali’, Pyrus ussuriensis Maxim.; and ‘Chuwhangbae’, ‘Wonwhang’, and ‘Imamuraaki’, Pyrus pyrifolia Nakai. For electron microscope imaging, pollen frozen at - 20°C was put on wet filter paper placed in 300-mL beakers and treated in a cold room at 5°C for the moist-chamber process for about 6 hours before use. A scanning electron microscope (SEM, S-3500N, Hitachi, Japan) was utilized under low vacuum (100 Pa) without coating, and the samples were observed at an acceleration voltage of 10 kV. The whole pollen forms were imaged at 500× magnification; one particle of pollen was images at 2,000× magnification; and the pollen surface pattern was imaged at 15,000× magnification to measure the size.
To investigate the morphological characteristics of pollen, the P/E ratio, the ratio of polar to equatorial diameters, was measured and compared among the varieties according to pollen forms. For further examination, they were imaged with a scanning electron microscope at 15,000× magnification to determine the widths and distance between striate bumps in the pollen surface patterns and pore distribution status in the lireas between striate bumps.
Pollen Germination Rate
For this experiment, pollen from the same five varieties used in the pollen morphologic characteristic investigation was employed. Filter paper was placed on 300-mL beakers and soaked with distilled water. Pollen kept in a freezer at - 20°C was put on the paper in the beakers and treated in a cold room at 5°C for the moist-chamber process for about 6 hours before use. For the pollen germination experiment, preformulated medium [1% agar (Daejung, Korea), 10% sucrose (Biosesang, Korea), and 200 ppm of boric acid (Yakuri Pure Chemicals Co., Ltd., Japan) mixed into 100 mL of water] was placed on slide glasses as thin as about 1 mL using a pipette. A specify amount of pollen was placed on a cotton swab and treated in a cold room at 5°C for the moist-chamber process for about 6 hours. The pollen was dropped evenly on the medium at about 50 cm distance between the swab and the slide. Filter paper was placed on the slide glasses and soaked with distilled water. Parafilm (PM-996, Bemis, USA) was used to seal the pollen inside petri dishes and a constant temperature was maintained. The samples were placed in a pyrostat at 25°C (DS-14MC, Dasol Scientific, Korea) to observe the pollen germination status every 2 hours for up to 12 hours.
Pollen germination rate was investigated using a stereomicroscope (MZ16A, Leica, Germany) at 50× magnification. Pollen tube elongation exceeding pollen diameter was determined as germination. The process was repeated three times for each variety, and three slide glasses were utilized in each repetition. Each slide glass was viewed five times, and 100 particles of pollen were investigated for their germination status in each view. The results were expressed as the germination rate (number of germinated pollen particles/number of total pollen particles × 100).
Pollen Tube Elongation
The same pollen varieties were utilized for the pollen tube elongation experiment (as the germination rate experiment above).
Filter paper was placed on petri dishes (90 × 15 mm, SPL Life Sciences Co., Ltd., Korea) and fully wet with distilled water. Pollen kept in a freezer at - 20°C was put on the petri dishes and treated in a cold room at 5°C for 3, 6, and 9 hours, respectively, for the moist-chamber process before use. For this experiment, placement was implemented in the same manner as that in the pollen germination experiment. The slide glasses with samples were then put in petri dishes on with filter paper fully wet in distilled water. To keep the pollen pollination and temperature constant inside the petri dishes, they were sealed using parafilm (PM-996, Bemis, USA) and then put in a culture medium (DS-14MC, Dasol Scientific, Korea) at 25°C and cultivated for 10 hours. After that, pollen tube elongation status was observed using a stereomicroscope (MZ16A, Leica, Germany) at 50× magnification. To examine pollen tube elongation, 100 single views were analyzed using the Leica Application Suite V4 program. Three moist-chamber processes were repeated three times hourly using three slide glasses in each round of repetition. Three views were investigated per slide glass.
Pollen Fertilization Rate and Fruit Weight
To examine the fertilization rate of pollen for pear pollination, five varieties of ‘Xuehuali’, ‘Yali’, ‘Chuwhangbae’, ‘Imamuraaki’, and ‘Wonwhang’ were studied in an orchard in Anseong-si, Gyeonggi-do, where 17-year-old ‘Niitaka’ pear varieties were planted.
For the preparation of pollen for pollination, filter paper were placed on 300-mL beakers and wet fully with distilled water. On the paper, the pollen of five varieties, which had been kept in a freezer at - 20°C, was taken out and put in the beakers then placed in a cold room at 5°C for about 6 hours for the moist-chamber process. The pollen processed in the moist chamber was put in 100-mL plastic bottles for use in pollination. One hundred pedicels per repetition were randomly placed on a glassine paper bag (10 × 5 cm) to block the flower-visiting insects 1 week prior to full bloom. On April 13, 2018, in the full bloom stage of the ‘Niitaka’ varieties, pollination was performed by putting the pollen on cotton swabs. After the artificial fertilization, the pollen was put back into the glassine paper bag.
Flowers labelled prior to fertilization were investigated. Fertilization rate was investigated by counting the number of fruit sets on May 2nd, 20 days after fertilization, and calculating the fertilization rate (number of fruit set/number of total flowers × 100). One hundred flowers were investigated a week in each round of repetition. Pear trees were cultivated and managed in accordance with general practices. On September 29th, the 189th day from full bloom, each process was repeated three times, and 30 fruits were randomly harvested in each round of repetition and weighed.
Statistical Analysis
For statistical analysis, a completely randomized design method was employed three times. Interprocess significance was analyzed at the level of Duncan’s multiple range test (p < 0.05) using SAS 9.4 (SAS Institute Inc., Cary, NC, USA).
Results and Discussion
Pollen Morphological Characteristics
The characteristics of five pollen varieties for pear pollination, ‘Xuehuali’, ‘Yali’, ‘Chuwhangbae’, ‘Imamuraaki’, and ‘Wonwhang’, were examined. The results are shown in Table 1 and Fig. 1. In terms of pollen size and form, the varieties were mostly prolate and perprolate types. ‘Xuehuali’, ‘Yali’, ‘Chuwhangbae’, and ‘Imamuraaki’ had P/E ratios between 1.87 and 1.98, belonging to the prolate type. ‘Wonwhang’ had a P/E ratio of 2.20, which was considered to be the perprolate type. Pollen mostly consists of a single monad. Some pollen consists of dyads, tetrads, or polyads, but the pollen in this experiment was monad (Lee, 1986). In general, pollen has diverse sizes with diameters ranging from 15 to - 300 µm. Flower size has a positive correlation with pollen size and is closely related to pollination methods. The size of the pollen fertilizable by wind is known to stay within 15 to - 45 µm and that of pollen fertilizable by insects is 15 to - 120 µm in diameter (Lee, 1986). Pear trees fertilize by wind and insects in natural conditions. The pollen diameters in this experiment were 21.20 to - 24.10 µm within this range (Table 1).
Table 1. Pollen size of the five varieties used in this study
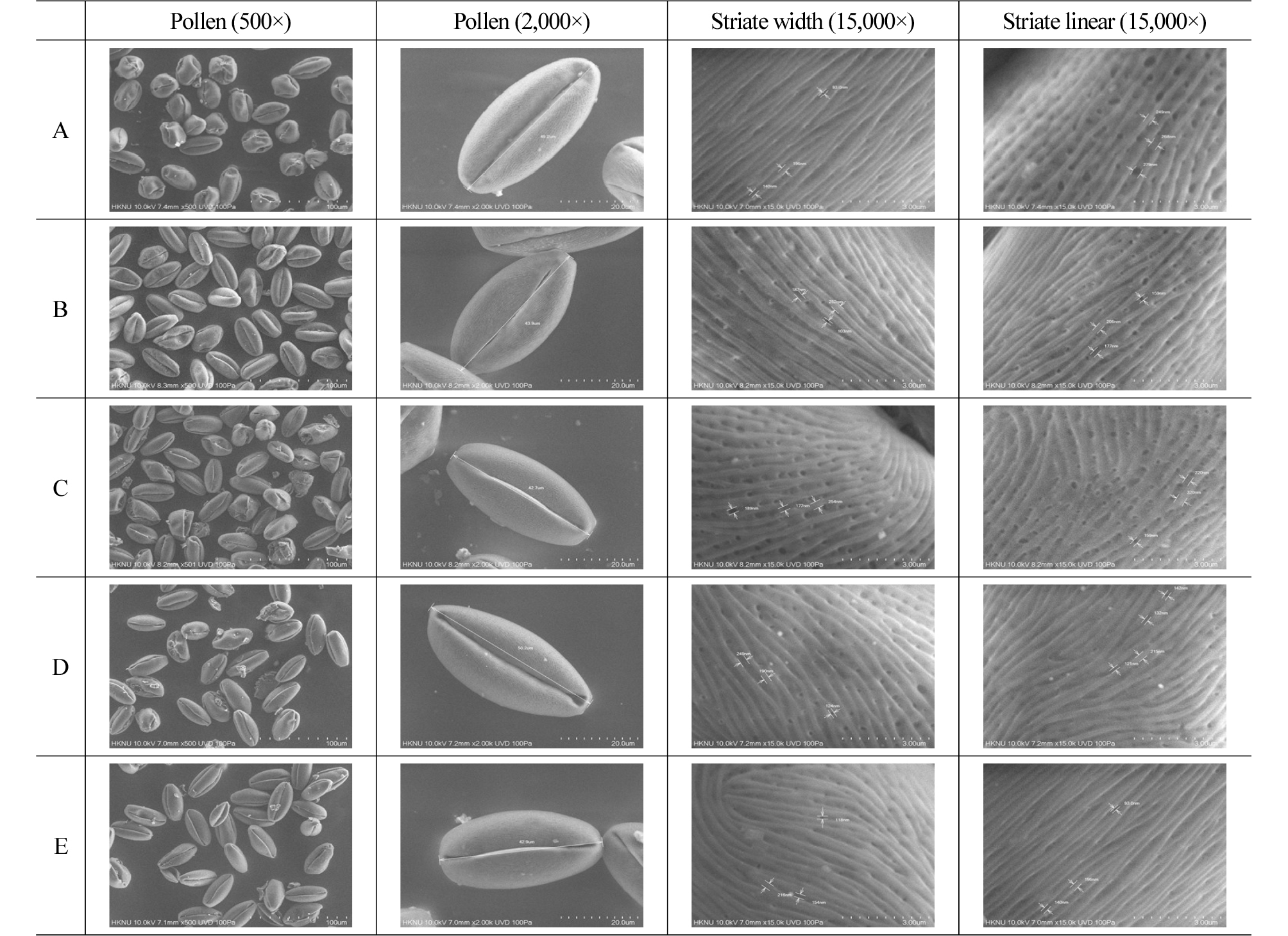
Generally, the types of pollen surface patterns are expressed in 12 categories, including psilate, pilate, and rugulate (Lee, 1986). ‘Xuehuali’, ‘Yali’, ‘Chuwhangbae’, ‘Imamuraaki’, and ‘Wonwhang’ were observed to have the striate type (Fig. 1).
In palynology, it is known that varieties can be identified by looking at their pollen surface patterns (Davis and Heywood, 1973; Lee, 1984). With respect to pear, Japanese pear and Chinese pear can be differentiated, but their varieties are hard to identify. It is reported that their varieties can be identified according to the size and frequency of pores in striate lireas (Shim et al., 1988b). Table 2 shows the striate bump width and striate linear width in the central part of pollen surface of each variety. The striate bump width of ‘Xuehuali’ was 249.8 ± 28.98 nm and that of ‘Imamuraaki’ was 230.6 ± 34.71 nm, which was wider. ‘Yali’, ‘Chuwhangbae’, and ‘Wonwhang’ showed narrower widths, but the differences were not statistically significant (p < 0.05).
Table 2. Morphological characteristics of the pollen surface of each variety for pear pollination
‘Xuehuali’, ‘Yali’, and ‘Imamuraaki’ had narrower ridge width than striate bump width, whereas ‘Chuwhangbae’ and ‘Wonwhang’ had wider ridge width than striate bump width. ‘Xuehuali’, ‘Yali’, and ‘Chuwhangbae’ had a deep valley of bumps, large pores, and high pore frequency (Fig. 1). However, ‘Wonwhang’ and ‘Imamuraaki’ had a lower valley of bumps and lower pore frequency. Based on these results, it seems possible to identify pollen varieties for artificial pollination by looking at their morphological characteristics, such as striate bump, valley depth, and pore frequency. Further detailed study would be necessary in this regard. However, in this study, we tried to see whether the characteristics of the pollen varieties were different from those of the varieties. Therefore, it is necessary to study morphological details of pollen varieties.
Pollen Germination and Pollen Tube Elongation
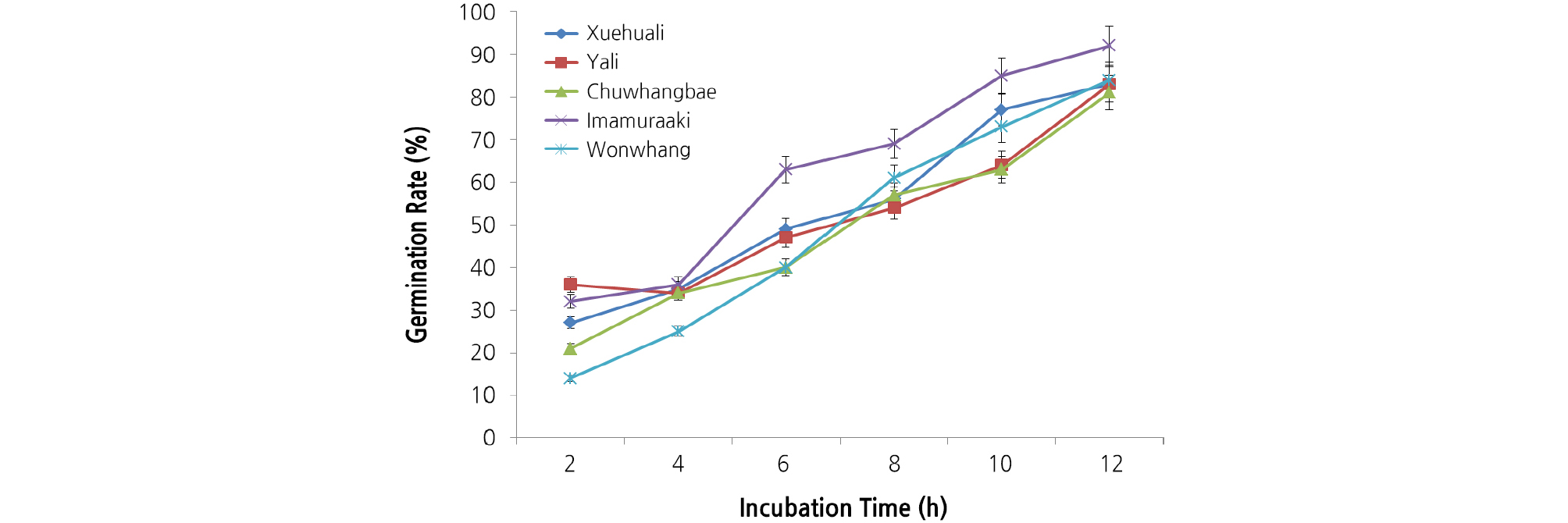
‘Xuehuali’, ‘Yali’, ‘Chuwhangbae’, ‘Imamuraaki’, and ‘Wonwhang’ pollen for pear pollination was investigated for germination rate. The results are exhibited in Fig. 2. Germination rate was observed for up to 12 hours from pollen placement every 2 hours. It was as low as 20 to 30% until 2 hours from pollen placement, while it rose to as high as 80% or over in all varieties after 12 hours (Fig. 2). Of all the varieties, ‘Imamuraaki’ was the highest at 92%. ‘Imamuraaki’ pollen had the most complete form among the five. ‘Imamuraaki’ also showed the best results in pollen germination rate, germination tube elongation, and seed weight (Peak and Cheong, 1977). The pollen used in this study was excellent regardless of the variety. General causes of poor pear fruit load include faulty fertilization due to restricted pollen germination by agricultural chemicals sprayed on pears during the blooming season (Peak and Cheong, 1977; Butt et al., 1985), pistil damage caused by low temperature led to no fruit set or fruit deformation (Kwon, 2006), and poor fertilization due to problems in the available pollen, such as purity, variety, cultivation environment, disease, and insects. Poor fruit loading due to faulty fertilization damages pear orchards and its causes have been analyzed in diverse aspects. We implemented a germination test to see if there was any problem with the pollen used for pollination and found that germination rate was at least 80%, indicating that the presently available five varieties are normal.
For pollination, it is a common practice to keep pear pollen in a freezer at around - 20°C, and carry out artificial pollination at room temperature in a wet manner. In this experiment, the wet process was applied for 3, 6, and 9 hours and then the pollen tube elongation status was investigated. After placing pollen in medium, it was cultivated for 10 hours. The pollen tube elongation results are in Table 3 and Fig. 3. After a 3-hour moist-chamber process, pollen tube elongation of ‘Xuehuali’ was 0.73 ± 0.06 mm, that of ‘Yali’ was 0.81 ± 0.04 mm, that of ‘Chuwhangbae’ was 0.63 ± 0.08 mm, and that of ‘Imamuraaki’ was 0.94 ± 0.15 mm, showing better performance than pollen subjected to 6- and 9-hour moist-chamber processes. However, ‘Wonwhang’ showed the best result of 1.32 ± 0.44 mm in the 9-hour moist-chamber processed group. These factors are thought to cause a decrease in the vigor of the pollen during the moist-chamber processes, but the pollen was good at 9 hours in ‘Wonwhang’ and required detailed study on the vigor of the pollen. Therefore, there seems to be not a big problem if frozen pollen is processed in a moist chamber for 3 hours. Spraying agricultural chemicals during the blooming season is considered a major factor influencing pollen tube elongation, but there are differences according to varieties (Kang et al., 2010). Given that varieties with good pollen germination and pollen tube elongation also have good fertilization and fruit loads, it is known that laboratory investigation and field experiment are correlated (Church and Williams, 1977). Chemical treatment is reported to disturb pollen tube elongation, which leads to low fruit-load rate, causing economic damage (Kang et al., 2010; Jung et al., 2017). Therefore, it seems that the varieties for pollination, which have a high pollen germination rate and good pollen tube elongation, lead to high fertilization rate and fruit load in the farms.
Table 3. Pollen tube elongation for artificial pollination after 10-hour cultivation from moist-chamber treatment

Fruit-Set Rate and Fruit Weight of Pollen for Pollination
‘Niitaka’ stigmas were pollinated with the pollen of five varieties of ‘Xuehuali’, ‘Yali’, ‘Chuwhangbae’, ‘Imamuraaki’, and ‘Wonwhang’ using cotton swabs. Their fruit-set rates were investigated 20 days from the pollination. As a result (Table 4), ‘Imamuraaki’ and ‘Wonwhang’ showed 97.0 and 96.3%, the highest levels and the other three varieties showed fertilization rates ranging from 92.3 to 94.6%. Thus, it was found that the pollen of five varieties utilized in this experiment was functional for pollination. Pear weight was investigated during the harvest period. As a result (Table 4), pollen varieties showed differences ranging from 580.3 to 611.8 g but with no statistically significant difference (p < 0.05). Pollen from each variety had a positive correlation between fruit-set rate and pear weight (R2 = 0.9288) and the higher the fruit-set rate, the heavier the pear weight was. With respect to the influence of pollen on fruit form, such an effect was reported different according to the number of seeds per piece of fruit (Tufts and Hansen, 1933). Shin et al. (1988) reported that metaxenia could not be recognized in pears. In this experiment, the ‘Niitaka’ variety was artificially pollinated with the pollen of five varieties, and it was found to have no effect on the size of pears.
Table 4. Fruit weight at harvest time and fruit-set rate 20 days after artificial pollination of 'Niitaka' variety with pollen from the five varieties in this study
| Pollen varieties | Fruit set (%) | Fruit weight (g) |
| Xuehuali | 92.6 bz | 580.3 a |
| Yali | 92.3 b | 585.2 a |
| Chuwhangbae | 94.6 b | 601.8 a |
| Imamuraaki | 97.0 a | 611.8 a |
| Wonwhang | 96.3 a | 605.4 a |
