

서 언
재료 및 방법
식물재료 및 재배조건
생육특성 조사
광반응 및 CO2 반응 곡선의 분석
엽온별 광합성률과 호흡률의 측정
통계분석
결 과
환경요인의 변화
마늘 지상부의 외형적 특성
광량 및 CO2 농도 변화에 대한 광합성 반응
엽온에 따른 광합성의 변화
마늘의 생산성과 품질
고 찰
서 언
마늘(Allium sativum L.)은 백합과(Liliaceae) 파속(Allium)에 속하는 초본성 작물로 유럽, 아시아, 아 메리카 대륙 등 세계 각지에서 재배되고 있으며, 우리나라에서는 중부와 남부 지방에서 재배되고 있다. 마늘은 기후 특성에 따라 분포와 생육의 뚜렷한 차이를 보이며, 기후는 생산량, 재배 기간 등에 다양하게 영향을 미친다(Cortés et al., 2003). 마늘은 인편이나 주아 등 영양번식에 의해 증식되므로 재배되는 각 지역의 환경에 적응되어 있으며, 생육지역의 기후 특성에 따라 한지형 마늘과 난지형 마늘로 구분하고 있다(Hwang and Lee, 1990; Hwang et al., 2009). 한지형 마늘은 의 성, 단양 등 비교적 기온이 낮은 중부지방을 중심으로 재배되는 데 반해, 난지형 마늘은 기온이 높은 신안, 무안, 해남, 고흥, 남 해 등 남해안 일대와 제주도에 주로 재배되고 있으며 국내 마늘 재배면적의 80%를 차지한다(Lee et al., 2005; Nam et al., 2008).
마늘은 생육초기에 일정기간 저온기를 거친 후 고온과 장일조건 하에서 인경이 비대하는 특성을 가지고 있어서(Bandara et al., 2000; Rahim and Fordham, 2001), 우리나라에서는 가을에 파종하여 겨울을 거친 후 이듬해 초여름에 수확을 하고 있다 (Song et al., 2001). 따라서 한지형 마늘을 난지에 재배하면 저온 요구도를 충족시키지 못하여 인경이 충실히 비대하지 못하 고, 반대로 난지형 마늘을 한지에 재배하면 겨울 동안에 동해를 받아 맹아의 출현율이 낮을 뿐만 아니라 월동 후 생육이 지연되 어 수량이 감소된다(Ogawa et al., 1975). 이처럼 마늘은 생육기간이 길고 주로 노지에서 재배되기 때문에 주위 환경에 많은 영향을 받으며, 특히 월동 전후 및 구 비대기의 기상조건에 따라 매년 수확량이 크게 달라질 수 있다. 한지형 마늘은 낮은 주야 간 온도(14/10 - 17/12°C)에서 재배하였을 때 인경 당 정상적인 인편 발생률이 90% 이상이고 균일한 크기와 중량의 인편들이 형성되지만, 온도가 높아짐에 따라 소인편 발생과 이차생장에 의해 상품성이 저하되고, 높은 주야간 온도(28/23°C)에서는 인 편 분화가 전혀 이루어지지 않아 통구를 형성한다(Oh et al., 2017).
본 연구에서는 난지형 마늘의 재배에 있어서 적합한 재배온도 범위를 알아보기 위하여, 주야간 온도(11/7 , 17/12, 23/18°C)를 달리한 soil-plant-atmosphere-research(SPAR) chamber에서 마늘을 키우면서 저온과 고온 스트레스에 의한 생육 특성, 광합성, 수확기 인편 특성 등의 변화를 살펴보았다.
재료 및 방법
식물재료 및 재배조건
본 실험에 사용한 마늘(Allium sativum L.)은 난지형인 ‘남도(Namdo)’이며, 5 - 6g 정도 되는 중간 크기의 건전한 인편을 종 구로 사용하였다. 10월 초순에 soil-plant-atmosphere-research(SPAR) 체임버 내부의 토양에 3열로 20 × 15cm 간격으로 인 편을 파종하고 충분히 관수한 후, 1주간 체임버를 개방하여 자연조건에서 발아시켰다. 이후 주야간 재배온도를 11/7, 17/12, 23/18°C로 일정하게 유지하도록 복합환경제어시스템(TGC-Soldan, Soldan Crop., Korea)을 이용하여 제어하여 2014년 10 월 17일부터 이듬해 5월 15일까지 재배하였다.
SPAR 체임버(SPAR facility, USDA-ARS, MS, USA)는 농촌진흥청 온난화대응농업연구소(N33°28' E126°31', 200m above sea level, Jeju, Korea)의 시설을 이용하였으며, 각각의 온도 별로 3열로 2개의 체임버를 사용하였다. 체임버는 2.2m × 1.4m × 2.5m (length × width × height)이며, 1.27cm 두께의 플렉시유리(plexiglass)로 둘러싸여 있어 재배기간 동안 97% 이 상의 자연광을 이용할 수 있도록 제작되었다(Reddy et al., 2001). 체임버 내부의 주야간 온도는 설정온도 ± 0.5°C 범위가 되도 록 조절하였으며, 한국천문연구원 월별, 지역별 해/달 출몰시각을 토대로 일출시각부터 주간 온도로, 일몰시각부터 야간 온도 로 1주일 단위로 전환되도록 설정하였다.
체임버 내 대기 온도와 일사량은 지상부로부터 2.5m 높이에 설치한 온도센서(1400-101, LI-COR Inc., USA)와 광센서(LI200X, LI-COR Inc., USA)를 이용해 측정하고, 데이터로거(CR1000, Campbell Scientific Inc., USA)에 연결하여 1시간 간격으로 기록하였다. 체임버 내 CO2 농도는 400µmol·mol-1로 유지되도록 조절하였다. 토양수분은 재배기간 동안 3일 간격 으로 충분히 관수하였으며, 기타 재배관리는 농촌진흥청 표준영농재배법에 준하여 관리하였다.
생육특성 조사
마늘 지상부의 외형적 특성은 온도 처리 후 20일(늦가을, 11월 초순)부터 210일(늦은 봄, 5월 중순)까지 7일 간격으로 10개 체를 무작위로 선정하여 지상부 길이, 엽초경, 엽수 등을 조사하였다. 지상부의 길이는 잎을 길게 늘어뜨렸을 때 인경 바로 윗부 분에서부터 잎의 끝부분까지의 길이를, 엽초경은 맹아엽 하단부의 직경을 측정하였다. 엽수는 맹아엽을 제외한 주당 모든 엽 수로 나타내었다. 총포 너비, 인경 특성(인경경, 인경고, 무게, 인편수 등), 인편 특성(인편의 너비, 길이, 무게 등) 등은 수확기 (DAT 210, 5월 중순)에 조사하였다.
광반응 및 CO2 반응 곡선의 분석
광반응 곡선(A-PPFD curves)은 온도 처리 후 150일째(3월 중순)의 식물체 4 - 5개체를 대상으로 LED 광원이 부착된 휴대형 광합성 측정기(LCpro+ Portable Photosynthesis System, ADC Bio Scientific Ltd., Hoddesdon, UK)를 이용하여 광량(0, 25, 50, 100, 200, 400, 800, 1,200, 1,600, 1,800µmol·m-2·s-1)의 변화에 대한 광합성 반응을 측정하여 작성하였으며, 이로부터 최대광합성률(Amax), 암호흡률(Rd), 광보상점(Qcomp), 광포화점(Qsat), 반포화상수(K), 순양자수율(φ)을 산출하였다(Marshall and Biscoe, 1980; Oh et al., 2015).
CO2 반응 곡선(A-Ci curves) 또한 온도 처리 후 150일째(3월 중순)의 식물체 4 - 5개체를 대상으로 LED 광원과 CO2 분사시 스템이 부착된 휴대형 광합성 측정기(Li-6400 Portable Photosynthesis System, Li-Cor Biosciences, Lincoln, NE, USA)를 사용하여 엽육 내 CO2 농도(25, 50, 100, 200, 300, 400, 600, 800, 1,200µmol·mol-1)를 달리하여 광합성량을 측정하여 작성 하였으며, 이를 토대로 최대전자전달률(Jmax), 광호흡률(Rp), 최대카르복실화속도(Vcmax), CO2 보상점(CCP), CO2 포화점 (CSP), 탄소고정효율(ACE)을 산출하였다(Farquhar et al., 1980). 광합성 반응 측정 시에 leaf 체임버는 20 ± 1°C, 1,200 µmol·m-2·s-1의 포화광을 조사하였다.
광반응 곡선과 CO2반응 곡선은 SigmaPlot 10.0 (Systat Software Inc., Erkrath, Germany)을 이용하여 작성하였으며, non-rectangular hyperbola equation으로 나타내었다.
엽온별 광합성률과 호흡률의 측정
엽온별 광합성률과 호흡률은 휴대형 광합성 측정기(LCpro+ Portable Photosynthesis System, ADC Bio Scientific Ltd., Hoddesdon, UK)를 이용하여 엽온을 15 - 40°C로 5°C 단위로 증가시켜 엽온의 상승에 대한 광합성과 호흡의 변화를 조사하 였다. 광합성률(photosynthetic rate, A)은 잎 챔버 내 CO2 농도가 대기의 CO2 농도와 비슷해지면 마늘의 잎을 챔버에 물리고 5분간 안정화시키고, 각 설정 엽온 하에서 1,200µmol·m-2·s-1의 포화광을 비추어 측정하였다. 호흡률(dark respiration, Rd)은 광합성률과 같은 방법으로 측정하였으며, 다만 포화광을 비추는 대신에 암조건을 유지하였다.
통계분석
마늘의 생육 특성과 광합성 특성에 대한 통계분석은 SPSS 통계 패키지 18.0(SPSS, Chicago, IL, USA)를 이용하여 수행하였으며, ANOVA에 이은 Duncan의 다중검정(p < 0.05)으로 평균치 간의 차이에 대한 유의성을 검정하였다.
결 과
환경요인의 변화
실험기간 동안 외부 기상 환경요인으로 일장과 매일 적산일사량의 변화를 살펴보았다(Fig. 1). 일장은 10월 중순 이후 12월 중순까지 감소하다가 이후에는 점차 길어졌으며 3월 중순부터는 12시간 이상으로 길어졌다(Fig. 1A). 매일 적산일사량은 기 복이 심하였으나, 10월 중순부터 점차 감소하다가 12월을 기점으로 다시 증가하는 양상을 보였다. 특히, 매일 적산일사량이 12 월에 가장 낮았으며 12월과 1월에 10MJ·m-2 이하인 날이 많았고 3월 이후에는 20MJ·m-2를 상회하였다(Fig. 1B). 체임버 내부 의 기온은 설정한 온도와 유사하게 유지되었으며(Fig. 1C), 각 체임버의 평균온도는 7.8, 14.0, 18.7°C로 나타났다(Table 1). 체임버 내 토양의 평균온도는 각각 9.0, 13.5, 17.4°C로 대기 온도와 유사하거나 또는 다소 낮거나 높게 유지되었다(Table 1).
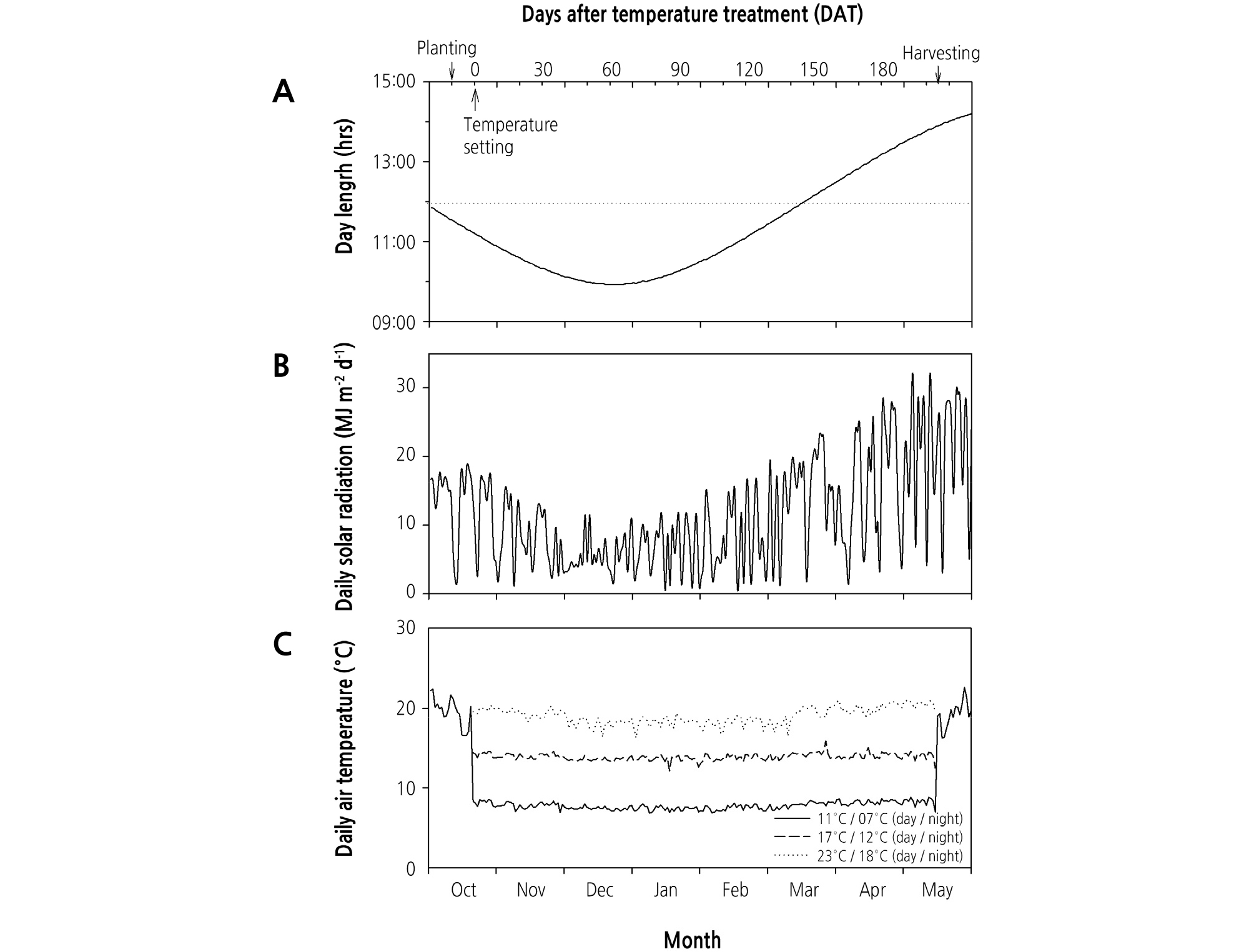
Table 1. The air and soil temperature in the SPAR space during the experimental period
yThe values within a column followed by different letter represent significant difference between setting temperatures.
마늘 지상부의 외형적 특성
생육 초·중기 (DAT 30 - 150)에는 모든 온도처리에서 초장이 길어지고, 엽초경이 굵어지고, 엽수가 증가하였으며, 고온 조건이 지상부 생육에 우호적으로 작용하였다(Fig. 2). 다만 엽초경은 주야간 17/12°C와 23/18°C에서 거의 같은 양상을 보였다 (Fig. 2B). 생육 후기 (DAT 150 이후)에는 주야간 17/12°C와 23/18°C에서는 지상부 생장이 거의 멈추고 엽초경이 점차 가늘 어지고 엽수도 급격하게 감소하였다. 주야간 23/18°C에서 재배하였을 때 엽초경과 엽수의 감소는 각각 170일과 140일 이후 인 데 반해 17/12°C에서는 각각 180일과 160일 이후로 늦춰졌다. 반면에 주야간 11/7°C에서 재배하였을 때에는 150일 이후 수확 시까지 지속적으로 초장이 길어지고 엽초경도 굵어지고 엽수도 증가하는 등 영양생장이 늦게까지 이루어졌으며, 190일 이후에야 엽수가 감소하였다(Fig. 2). 추대기는 재배 온도별로 차이를 보여 주야간 23/18°C에서 빠르고, 주야간 11/7°C에서 늦게 나타났다. 즉, 주야간 23/18°C에서는 3월 중순에, 주야간 17/12°C에서는 4월 중순에, 그리고 주야간 11/7°C에서는 5월 초순에 각각 화경이 발달하여 재배 온도 간에 대략 50일 정도 차이가 나타났으며(Fig. 2C), 수확기에 주야간 23/18°C와 17/12°C에서는 화경의 정단부에 성숙한 상태의 총포를 관찰할 수 있었으나, 주야간 11/7°C에서는 미숙한 상태였다.
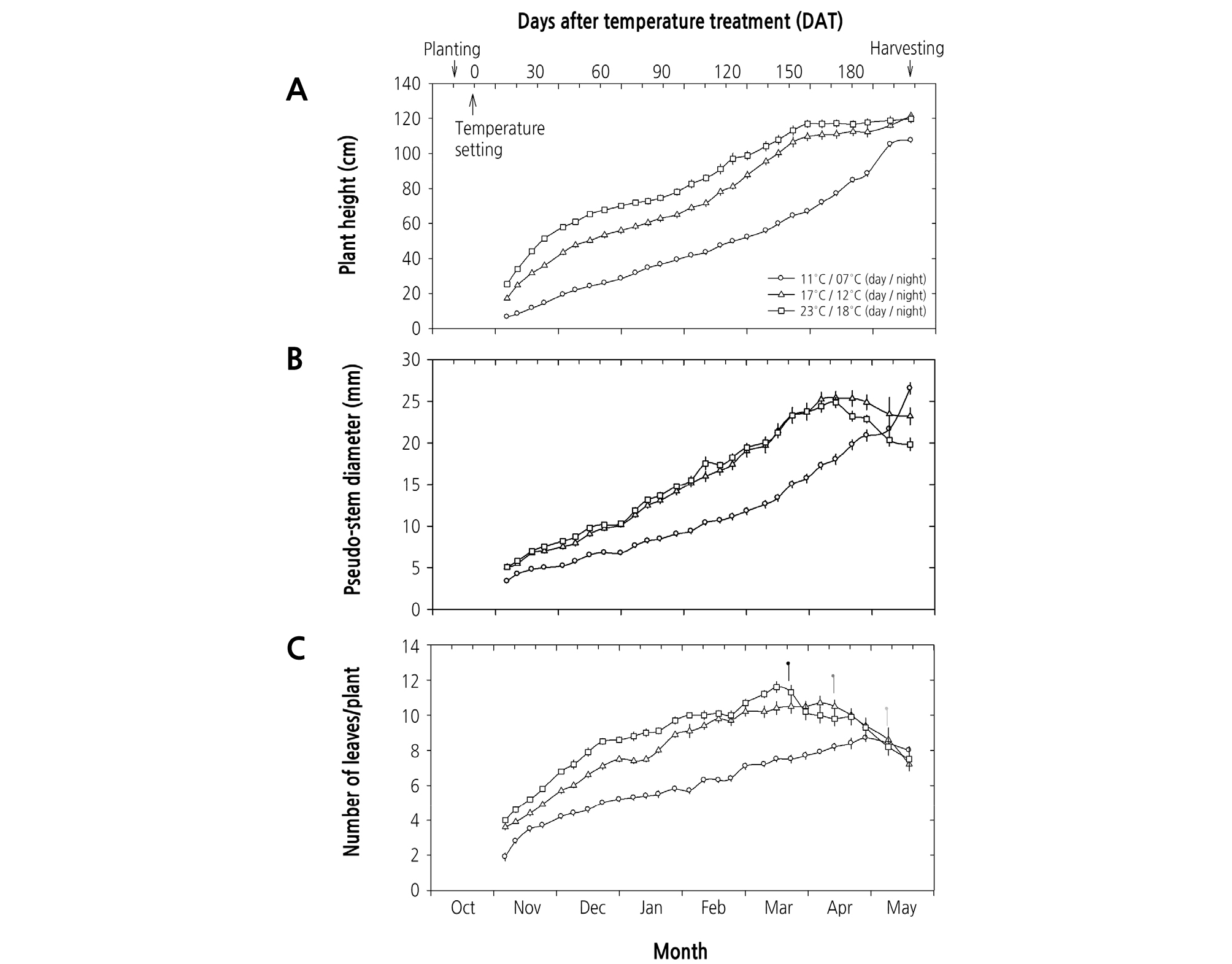
Fig. 2.
Effect of day/night temperatures on growth characteristics (A, plant height; B, pseudo-stem diameter; C, number of leaves/plant) of garlic (Allium sativum L.) during the experimental period. The upper marks of C represent the bolting times under each temperature set. Each value represents the mean ± SE measured from 10 independent plants.
광량 및 CO2 농도 변화에 대한 광합성 반응
마늘 잎의 광반응 곡선(A-PPFD curve)과 CO2 반응 곡선(A-Ci curve)을 작성하고(Fig. 3), 이로부터 여러 가지 광합성 특성을 조사하였다(Table 2). 광반응 곡선(Fig. 3A)으로부터 얻은 특성 중 최대광합성률(Amax)은 주야간 17/12°C에서 재배하였을 때 18.8µmol CO2·m-2·s-1로 높았으며, 11/7°C에서는 15.8µmol CO2·m-2·s-1로 낮았다. 호흡률(Rd)은 23/18°C와 11/7°C 조건 에서 재배하였을 때 각각 1.49µmol CO2·m-2·s-1와 1.28µmol CO2·m-2·s-1로 17/12°C조건에서의 0.63µmol CO2·m-2·s-1보다 2배 가량 더 높았다. 광보상점(Qcomp)과 광포화점(Qsat)은 23/18°C 조건에서 재배되었을 때 다소 높았으며, 반포화상수(K)와 양자수율(φ)은 재배 온도 간에 유의한 차이를 보이지 않았다. CO2 반응 곡선(Fig. 3B)으로부터 얻은 특성 중 최대전자전달률 (Jmax)은 23/18°C 조건에서 재배되었을 때 31.5µmol·m-2·s-1로 높았으며, 광호흡률(Rp)은 17/12°C에서 재배하였을 때 다소 높으나 통계적으로 유의한 차이는 보이지 않았다. 그리고 최대카르복실화속도(Vcmax), CO2 보상점(CCP)과 탄소고정효율 (ACE)도 17/12°C 조건에서 다소 높았으나 재배 온도 간에 통계학적으로 유의한 차이가 없었다. 그리고 CO2 포화점(CSP)은 23/18°C 조건에서 재배되었을 때 다소 높았으며, 17/12°C 조건에서 오히려 낮았다.
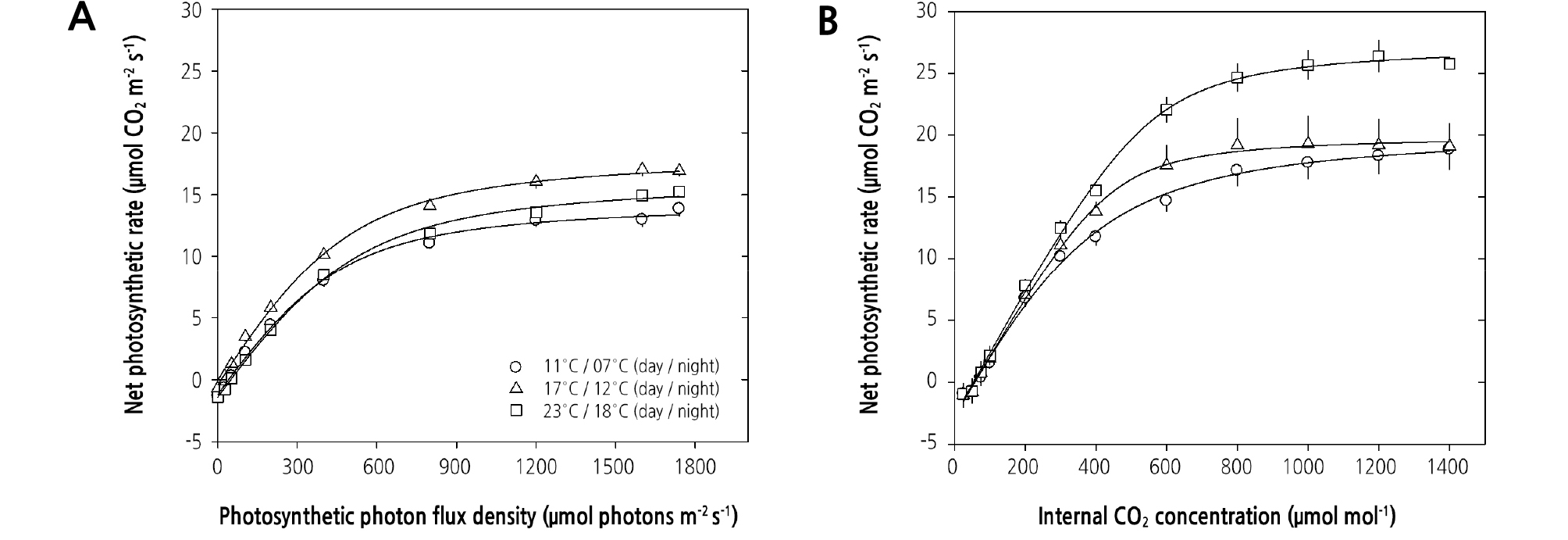
Fig. 3.
Photosynthetic light- response curves (A) and CO2-response curves (B) obtained from leaves of garlic (Allium sativum L.) grown under different day/night temperature regimes. The curves were fitted with the non-rectangular hyperbola. Each value represents the mean ± SE measured from 4 independent plants.
Table 2. Characteristics obtained from the photosynthetic light- and CO2-response curves in leaves of garlic (Allium sativum L.) grown under different day/night temperature regimes
yEach value represents the mean ± SE measured from four independent plants.
xDifferent letters in each row indicate significant difference among different day/night temperature regimes (p < 0.05).
엽온에 따른 광합성의 변화
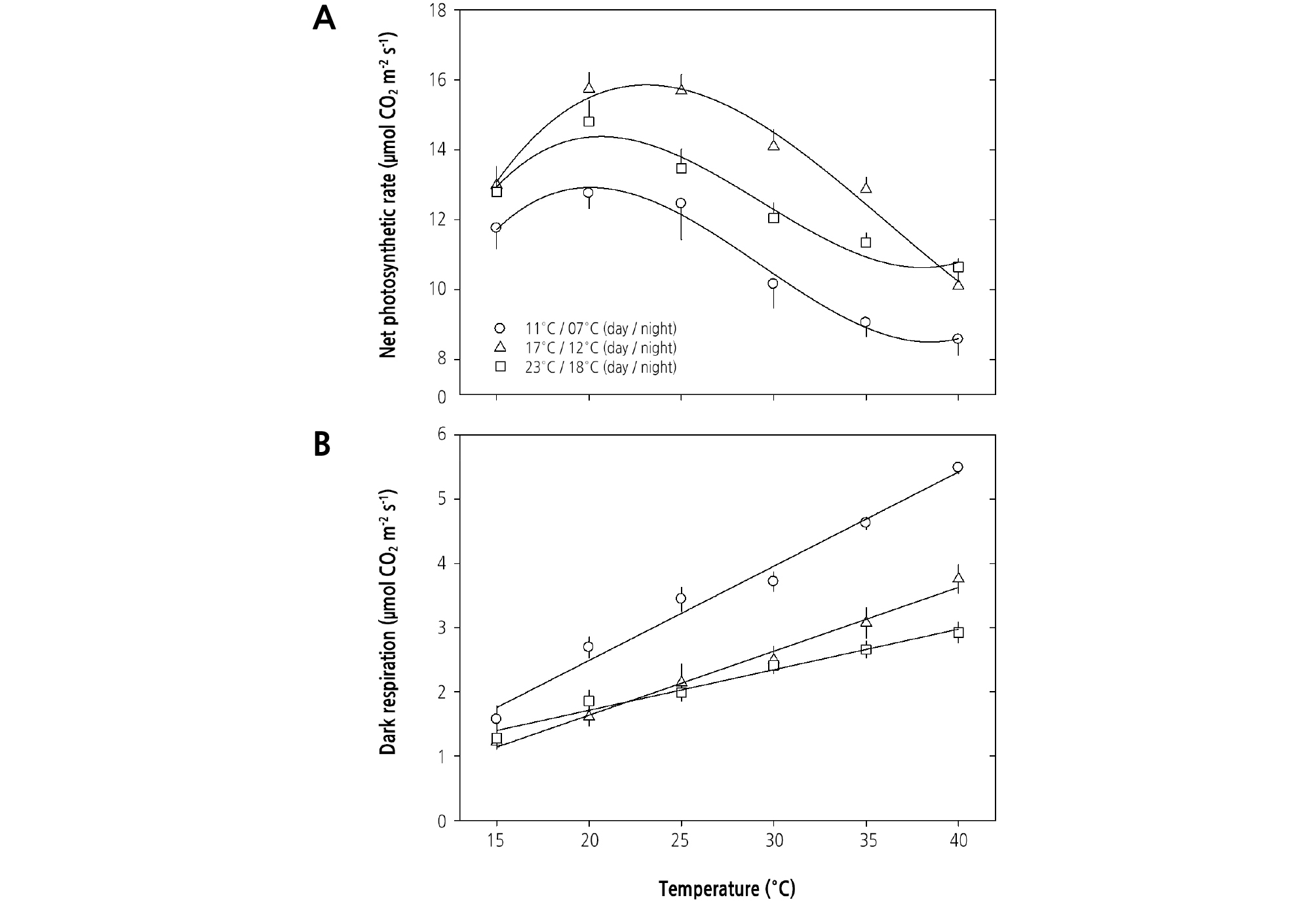
엽온별 광합성률은 재배온도에 따라 차이가 있지만 대략 20 - 25°C 범위에서 가장 높았으며, 암호흡률(Rd)은 모든 처리구에 서 온도 증가에 비례하여 증가하였으나 증가의 폭은 11/7°C에서 더 컸다(Fig. 4). 즉, 주야간 재배온도를 11/7°C, 17/12°C, 23/18°C로 조절된 조건에서 마늘을 재배하였을 때 광합성을 위한 최적 온도는 각각 20°C, 23°C, 21°C로 나타났으며, 이때 광합성률은 12.0µmol CO2·m-2·s-1, 15.0µmol CO2·m-2·s-1, 14.0µmol CO2·m-2·s-1였다(Fig. 4A). 반면에, 암호흡률(Rd)은 마늘이 자라는 재배온도에 관계없이 온도 증가에 비례하여 증가하였으며, 특히 주야간 11/7°C에서 재배하였을 때는 주야간 17/12°C와 23/18°C에서 재배하였을 때보다 더 컸다(Fig. 4B).
마늘의 생산성과 품질
수확기에 지상부는 23/18°C와 17/12°C에서 길게 신장하였으며, 엽초경은 11/7°C에서 굵고 재배온도가 높을수록 가늘어졌다. 엽수는 7 - 8개로 재배 온도 간에 통계적으로 유의한 차이는 보이지 않았다(Table 3). 화경의 기부에 형성된 총포는 재배 온도가 높을수록 빠르게 발달하여 두께가 굵어졌으며 지상부의 생체중은 재배 온도가 높을수록 감소하였다. 즉, 주야간 23/18°C와 17/12°C에서는 화경의 정단부에 성숙한 상태의 총포를 관찰할 수 있었으나, 주야간 11/7°C에서는 추대가 늦게 이 루어져 미숙한 상태였다.
Table 3. Growth characteristics of garlic (Allium sativum L.) grown for 210 days under different day/night temperature regimes
yDifferent letters in each row indicate significant difference among different day/night temperature regimes (p < 0.05).
그리고, 수확기에 각 재배 온도별 수확한 마늘의 외형적 특성을 살펴보면, 17/12°C에서 재배되었을 때에는 크기가 크고 상 품성이 높은 인경이 형성되었을 뿐만 아니라 인경 내 인편의 크기가 균일한 상태를 유지하였다. 주야간 11/7°C에서 재배되었 을 때에는 영양생장이 완료되지 않아 인경 크기가 작고 전체적으로 미숙한 인편이 형성되었으며, 23/18°C에서는 분화된 인편 속에 새로운 작은 인편들이 만들어져 벌마늘이 형성됨을 확인할 수 있다(Fig. 5). 수확기 인경 길이와 두께는 17 /12 - 23/18°C 에서 각각 3.6cm 이상과 7.0cm 이상으로 발달하였으며, 11/7°C에서는 인경 발달이 저조하였다. 그리고 인경 중량, 인편수와 인편 중량은 17/12°C에서 각각 120.6g, 8.4개, 9.7g으로 크기가 크고 상품성이 높은 인경이 형성되었다(Table 3). 그보다 높은 온도인 23/18°C에서는 인편 속에 새로운 작은 인편들이 분화되어, 즉 벌마늘이 형성되어 인편수가 19.0개로 2배 이상 증가하 였으며 인편 중량은 감소하였다. 인경당 정상적인 인편의 발생률은 17/12°C 이하의 온도에서 100%로 양호한 데 반해 23/18°C에서는 31.8%로 대부분이 소인편이 형성됨을 확인할 수 있었다.

고 찰
온도는 식물의 생장과 발달에 있어 중요한 환경인자로 생육적온을 벗어난 고온이나 저온은 작물의 생산성과도 직결되므로 고온 또는 저온 내성의 작물을 선발하거나 육성하는 작업들이 활발하게 진행되고 있다(Shu et al., 2016; Zhou et al., 2018). 마늘(Allium sativum L.)의 생장과 인경 형성은 생육 초기 일정기간의 저온기를 거친 후 고온과 장일 조건에서 이루어지므로 온도는 마늘의 생산에 있어서 중요한 환경요인이다(Takagi, 1990; Sohn et al., 2010).
재배온도별 마늘 잎의 광합성은 Fig. 3과 Table 2에서처럼 주야간 17/12°C에서 최대광합성률(Amax)은 높고, 암호흡률(Rd) 은 낮아 마늘재배에 최적인 것으로 보인다. 이는 Fig. 2의 재배기간별 지상부의 생육과 Table 3의 수확기 인경 크기와 중량, 인 편수와 인편 중량으로도 확인할 수 있다. 즉, 주야간 17/12°C에서 마늘을 재배하였을 때 생육기간 동안 지상부 신장과 엽수 증 가가 활발하고 수확기에 초장, 인경 두께와 길이, 인경 중량, 정상적인 인편수가 증가하여 상품성 높은 마늘을 수확할 수 있었 다. 이에 반해 주야간 23/18°C와 11/7°C에서는 최대광합성률(Amax)은 낮고, 암호흡률(Rd)은 높게 나타났다. 엽온별 광합성률 은 주야간 재배온도에 따라 차이가 있지만 대략 20 - 25°C 범위에서 가장 높았으며, 암호흡률(Rd)은 모든 처리구에서 온도 증 가에 비례하여 증가하였다(Fig. 4). 즉, Fig. 4A에서 살펴본 바와 같이 주야간 11/7°C, 17/12°C, 23/18°C로 조절된 조건에서 마늘을 재배하였을 때 광합성을 위한 최적의 엽온은 각각 20°C, 23°C, 21°C이며, 특히 주야간 17/12°C에서는 엽온별 광합성 률이 상대적으로 높았다. 한지형 마늘에서도 광계II의 최대 광화학적 효율(Fv/Fm)과 잠재적 광합성능(Fv/Fo)이 주야간 14/10 - 20/15°C 범위에서 가장 높고 지상부 생장과 인경 발달도 양호하여 마늘의 생육에 우호적이었으며, 20/15°C 이상의 온도에 서는 잎줄기가 터지거나 마늘통이 갈라지거나 벌마늘을 형성하는 등의 이차생장이 발생하였다(Oh et al., 2017). 호냉성 채소로 잘 알려진 배추(Brassica campestris L. ssp. napus var. pekinensis Makino)의 광합성은 15 - 30°C 범위에서는 엽온이 증가함에 따라 광합성 속도가 증가하나 30°C 이상의 온도에서는 감소하고, 광합성을 위한 최적의 온도가 26.1°C임이 보고된 바 있다(Oh et al., 2015). 그리고 Fig. 4B에서 살펴본 바와 같이 주야간 11/7°C, 17/12°C, 23/18°C에서 마늘을 재배하였을 때 호흡률(Rd)은 마늘이 자라는 재배온도에 관계없이 엽온 증가에 비례하여 증가하였다. 특히 주야간 17/12°C와 23/18°C에서 보 다 11/7°C에서 재배하였을 때 엽온 상승에 따른 암호흡률은 크게 증가하였는데, 이는 Fig. 2에서와 같이 11/7°C에서는 영양생 장이 완료되지 않아 호흡량이 증가하고, 그보다 높은 온도에서는 작물의 노화가 진행됨에 따라 점차 호흡량이 감소하고 지상 부의 생장률이 감소한 데에서 기인한 결과로 보인다.
Fig. 2에서와 같이 초장의 길이, 엽초경의 굵기나 엽수 등 지상부의 생육이 주야간 23/18°C와 17/12°C에서는 원활하게 이루 어져 수확기에 Fig. 5에서 살펴본 바와 같이 인경 형성에 있어서도 영향이 없으나, 주야간 11/7°C에서 재배하였을 때에는 생육 이 지연되어 수확기에도 영양생장이 지속적으로 이루어져 인경 비대가 거의 이루어지지 않았다. 마늘의 수확량은 인경 직경과 무게, 인편수 등과 관련이 있다(Panthee et al., 2006). 마늘의 인경 중량이 18 - 20°C 범위에서 최대값을 보이고 10°C 이하의 온도에서는 크게 낮아진다(Wi et al., 2017). 또한 온도구배터널을 이용하여 마늘을 고온조건에서 재배하면 외기온도에 비해 서 마늘 구의 무게는 크게 감소하지 않지만 외기온도 +6°C 고온구에서는 마늘 인편수가 증가하여 마늘 쪽당 무게가 감소하고, 벌마늘이나 스폰지마늘 등 비상품이 많아져 수량이 50% 정도 감소한다(Lee et al., 2018). 마늘의 생산성을 예측하는 데 있어 엽수는 중요한 요소이며, 특정 시점까지는 녹색 상태의 건전한 잎이 증가하고 이후 노화가 이루어지면서 점차적으로 녹색 상태의 잎이 감소하는 양상을 보인다(Moravčević et al., 2011; Oh et al., 2015). 본 연구에서도 Fig. 2C에서 살펴본 바와 같이 이러 한 특성을 잘 보여주고 있으며 최대 엽수의 시점은 23/18°C, 17/12°C, 11/7°C에서 각각 140일(12개), 160일(10개), 190일(8개)이며 그 이후에는 잎의 노화로 점차 감소하였고(Fig. 2C), 특히 저온에서 생산성이 낮음을 예측할 수 있다. 수확기에 엽수는 7 - 8개로 재배 온도 간에 통계적으로 유의한 차이는 보이지 않았으나, 이는 Fig. 2C에서 살펴본 바와 같이 17/12°C 이상의 온 도에서는 잎의 노화에 의해 녹색의 잎 수가 감소한 데 기인한 결과라고 할 수 있다.
한편, 인편의 발달에 있어서 고온과 장일 조건이 중요하게 작용하며(Kamenetsky et al., 2004), 인경 비대는 인편 분화에 관여하는 식물호르몬 등의 내적 요인이나 일장과 온도 등의 외적 요인에 의해서 영향을 받는 것으로 보고되고 있다(Takagi, 1990; Sohn et al., 2010). 인경 형성은 장일 하에서 촉진되며 12시간 이상의 일장에서는 완숙기가 단축되고 인경 수확량도 최 고에 달하게 된다. 일장과 일사량이 동일하였을 때, Fig. 5와 Table 3에서 보는 바와 같이 생육적온 내에서 재배온도가 높을수 록 수확기에 벌마늘 발생이 많아질 수 있으므로 23/18°C에서는 생육 후기에 고온 장해가 발생하지 않도록 수확시기를 다소 앞 당길 필요가 있다. 그리고 낮은 온도인 11/7°C에서는 영양생장이 완료되지 않아 미숙한 인편이 형성되었고 정상적인 인편을 얻기 위해서는 인경 비대가 충분히 이루어질 수 있도록 재배기간을 연장하여 수확시기를 늦출 필요가 있다. 난지형 마늘을 한 지에 재배하면 겨울 동안에 동해를 받아 맹아의 출현율이 낮을 뿐만 아니라 월동 후 생육이 지연되어 수량이 감소된다(Ogawa et al., 1975). 본 연구에서도 주야간 11/7°C인 저온에서는 생육이 지연되어 인경 비대 및 인편 형성에 영향을 미침을 확인할 수 있다. 마늘을 파종하였을 때 인편에서 맹아의 출현, 잎의 발달과 같은 초기 영양생장, 인경의 형성과 발달, 성숙은 마늘의 주요 발달 단계이며(del Pozo and González, 2005), 각각의 단계에 온도가 중요하게 작용한다. 한지형 마늘에서 이차생장 발생률 을 낮추면서 상품성이 높은 마늘을 생산하기 위해서는 재배기간 동안 14/10 - 17/12°C에서 재배하는 것이 바람직한 것으로 보고된 바 있다(Oh et al., 2017).
다양한 기상환경 중 온도는 작물의 생육에 가장 큰 영향을 주며, 생육 시기마다 알맞은 온도가 있고, 그 온도 범위를 벗어나면 저온 또는 고온에 의해 피해를 보게 된다. 본 연구에서는 주야간 17/12°C에서 재배하였을 때, 마늘의 최대광합성률(Amax)은 높 고 암호흡률(Rd)은 낮을 뿐만 아니라 크기와 모양이 균일한 인경과 인편을 생산할 수 있어, 상품성이 높은 난지형 마늘을 생산 하기 위해서는 재배기간 동안 17/12°C에서 유지하는 것이 바람직한 것으로 나타났다. 반면에, 23/18°C에서는 고온 장해가 발 생하고, 11/7°C에서는 인경 비대가 충분히 이루어지지 않아 상품성이 크게 떨어졌다. 그러나 최근 온난화 등으로 인한 이상기 후가 발생하고 있고, 미래에는 폭염이나 가뭄 등의 기상재해가 더욱 심해질 것으로 예측되는 바 상품성이 좋은 마늘을 수확하 기 위해서는 온도뿐만 아니라 가뭄 등의 영향을 최소화할 수 있는 방안을 모색할 필요가 있다.
