

Introduction
Materials and Methods
Plant Materials and Genomic DNA Extraction
Polymorphism Test of Microsatellite Marker Candidates
Genotyping by M13-tailed PCR
Data Analysis
Results and Discussion
Development of Polymorphic Microsatellite Markers
Identification of the Citrus Varieties Bred in Korea using Polymorphic Microsatellite Markers
Introduction
Citrus is a major fruit crop produced in tropical and subtropical regions around the world. Citrus annually produces 146 million tons of fruit from a cultivated area of 7.2 million hectares worldwide. Sweet orange cultivars account for approximately 60% of annual citrus production (FAO, 2018). In 2017, citrus produced over 0.6 million tons of fruit in the arable land of over 20,000 hectares in Korea. The entire production amounted to approximately 0.83 billion USD (www.krei.re.kr). Citrus fruit is an important source of nutrition for humans due to its high contents of vitamin C, citric acid, and other functional compounds, including citrus-specific flavonoids such as hesperidin and tangeretin (Benavente-García and Castillo, 2008; Iranshahi et al., 2015). Despite the tremendous extent and value of citrus production, most citrus varieties have been selected from natural hybridizations and somatic mutations (e.g., bud sport and nucellar mutants), not from systematic and targeted breeding programs (Talon and Gmitter, 2008). Citrus is considered the most difficult crop for developing new varieties through conventional breeding. Unfavorable factors for conventional breeding in citrus include large plant size, long juvenile stage, self- and cross-incompatibility, pollen and/or ovule sterility, high level of heterozygosity, and apomictic nucellar embryony (Donmez et al., 2013).
Microsatellites, also referred to as simple sequence repeats (SSRs), are highly informative DNA markers due to their high degree of polymorphism, codominant mode of inheritance, and genome-wide distribution (including organelle genomes). The International Union for the Protection of New Varieties of Plants (UPOV) highly recommends the use of molecular markers such as microsatellites to verify new varieties (UPOV, www.upov.int). Microsatellite markers have been widely used for various molecular genetic analyses, such as genetic diversity and population structure studies (Kim et al., 2016; Chung et al., 2019), construction of genetic linkage maps (Chang et al., 2018), identification of zygotic/ nucellar individuals (Woo et al., 2019), and cultivar/genotype identification (Kim et al., 2019).
In Korea, systematic citrus breeding programs have been in operation since the Citrus Research Institute, National Institute of Horticultural and Herbal Science, was established in 1992. More than 30 citrus varieties have been developed from bud sport mutants, nucellar mutants, and genetic hybridizations by breeders at governmental institutes and breeding companies as well as private breeders. Development of new citrus varieties will greatly increase in the near future due to citrus breeding programs in government and company sectors. However, reliable technologies for identifying Korean citrus varieties that protect breeders’ rights and guarantee the quality of nursery plants are not yet established. Therefore, in this study, we report the development of polymorphic microsatellite makers to identify citrus varieties bred in Korea.
Materials and Methods
Plant Materials and Genomic DNA Extraction
Leaf samples of citrus varieties used in this study were obtained from two public institutes in the Republic of Korea: Agricultural Research and Extension Services, Jeju Special Self-Governing Province, and Citrus Research Institute, National Institute of Horticultural & Herbal Science. Harvested leaf tissues were rinsed with running tap water and then stored at ‑ 80°C until use. Genomic DNA (gDNA) was isolated from the leaf tissue using the Biomedic® Plant gDNA Extraction Kit (Biomedic Co., Ltd., Bucheon, Korea). The isolated gDNA was quantified using the DeNovix DS-11+ spectrophotometer (DeNovix, Wilmington, DE, USA).
Polymorphism Test of Microsatellite Marker Candidates
Putative polymorphic microsatellite markers were selected from routine PCR using gDNAs from 11 varieties belonging to six citrus groups (lemon, mandarin, sour orange, citron, orange, and tangor) as described previously (Woo et al., 2019). The PCR reaction and cycling conditions were described in our previous report (Woo et al., 2019). PCR products were separated on a 2.5% (w/v) agarose gel to confirm PCR amplification and polymorphism among the tested varieties.
Genotyping by M13-tailed PCR
The M13-tailed PCR method was used for genotype analysis using the selected polymorphic microsatellite markers (Schuelke, 2000). The PCR reaction conditions were the same as those previously reported (Woo et al., 2019). The cycling conditions for PCR amplification followed the protocol described previously by Schuelke (2000). Fragment analysis of the PCR products was described previously (Kim et al., 2012). Calling of allele sizes was performed using GeneMapper software (ver. 4.0; Applied Biosystems).
Data Analysis
Genetic parameters such as major allele frequency (MAF), number of alleles (NA), genetic diversity (GD, often referred to as expected heterozygosity), observed heterozygosity (HO), and polymorphic information content (PIC) were measured by calculating the shared allele frequencies using PowerMarker software (v. 3.25) (Liu and Muse, 2005). The unweighted pair group method with arithmetic mean (UPGMA) dendrogram was implemented using MEGA software (v. 7.0) (Kumar, 2016), which is embedded in PowerMarker, with the UPGMA algorithm.
Results and Discussion
Development of Polymorphic Microsatellite Markers
We previously developed 17 polymorphic microsatellite markers and applied them both to analyze the genetic diversity of citrus genetic resources and to identify zygotic/nucellar individuals. However, the markers were not enough to discriminate all citrus genetic resources used in the study (Woo et al., 2019). Genome-wide characterization of microsatellite markers was reported in sweet orange (Citrus sinensis) previously (Biswas et al. 2014). We used marker information from the literature to further screen polymorphic microsatellites. To develop high-quality polymorphic markers, the microsatellite markers from sweet orange were applied to 11 citrus varieties belonging to six citrus groups: lemon, mandarin, sour orange, citron, orange, and tangor. We finally selected 36 polymorphic markers out of 74 microsatellite candidates. Table 1 summarizes 53 polymorphic nuclear microsatellite markers, including the 36 microsatellites developed in this study. The physical location of the 53 microsatellite loci on the nine pseudochromosomes was determined by performing a BlastN search against a reference genome of C. clementina cv. Clemenules (Wu et al., 2013; https://phytozome.jgi.doe.gov/pz/portal.html). The final microsatellite markers fully covered all of the pseudochromosomes of citrus (Fig. 1).
Table 1.
The 53 polymorphic microsatellite markers used in this study
| No. | Locus name | Primer sequence (5' → 3') | Repeat motif |
Tay (°C) |
Size range (bp) | Reference | |||
| Forwardz | Reverse | ||||||||
| 1 | BM-CiSSR-012 | TGTAAAACGACGGCCAGT GGGCTCAGTTCTTCTCTACTC | GCATTAGGCTTCTCTCATACC | (TTA)15 | 58 | 283-308 | Woo et al., 2019 | ||
| 2 | BM-CiSSR-013 | TGTAAAACGACGGCCAGT GGTGGCATACATACATACATACA | GCAACATCTGGAACTACTCA | (TA)6 | 58 | 127-135 | |||
| 3 | BM-CiSSR-032 | TGTAAAACGACGGCCAGT GCCTGAGTTTCTTTGTTATG | CATTCCATCGTCTCCTATTGT | (TATG)4 | 58 | 136-172 | |||
| 4 | BM-CiSSR-043 | TGTAAAACGACGGCCAGT ATTAGTGCGGGTAAGATGAA | AAGGATTTGGTGTAGGAAGTAA | (AAAAT)3 | 58 | 286-327 | Woo et al., 2019 | ||
| 5 | BM-CiSSR-073 | TGTAAAACGACGGCCAGT CGGACAAGGAGATGAAGATAG | TTCTAACAGCACCAAGCAG | (GA)16 | 58 | 319-345 | |||
| 6 | BM-CiSSR-077 | TGTAAAACGACGGCCAGT TGTATTTATTTCTGACTACGACC | ATGCGTTTGGTGTGTGTT | (AT)12 | 58 | 188-207 | |||
| 7 | BM-CiSSR-082 | TGTAAAACGACGGCCAGT ACCTGAGCCCTTTTTGGTTT | GCCAGATCAAGGCTCAAATC | (TC)13 | 58 | 150-155 | |||
| 8 | BM-CiSSR-087 | TGTAAAACGACGGCCAGT CAGATCCTATTGCAGAGGCA | GCCCATTTGTATTGCCATTT | (CAG)6 | 58 | 183-189 | |||
| 9 | BM-CiSSR-093 | TGTAAAACGACGGCCAGT CCCCCTCTTCTTTCACACAA | GGTGAGCAGCCATCTTCTTC | (TA)6 | 58 | 152-156 | |||
| 10 | BM-CiSSR-094 | TGTAAAACGACGGCCAGT GAATTGGGAGGACGAACTGA | CGAGCCCTAGACAGAGATGG | (AGA)7 | 58 | 268-277 | |||
| 11 | BM-CiSSR-100 | TGTAAAACGACGGCCAGT GTTTTCAGCTGGATTCGAGG | CACGTGTCCTCCTGGAACTT | (GCC)5 | 58 | 197-206 | |||
| 12 | BM-CiSSR-111 | TGTAAAACGACGGCCAGT CCGATACAGCACAAAGCAAA | TGGAAAGAGAGAAGCCAAGC | (AAT)7N15(AGC)7 | 58 | 134-143 | |||
| 13 | BM-CiSSR-115b | TGTAAAACGACGGCCAGT CGGTGTGTATTGGGTACACG | GCTTTTTCGAAAGCGTCAAG | (TA)17 | 58 | 231-250 | |||
| 14 | BM-CiSSR-137 | TGTAAAACGACGGCCAGT GCAACGTGTACTGACGCTTG | GCTCGTATCTGAAGCTCGCC | (TAT)7 | 53 | 301-327 | |||
| 15 | BM-CiSSR-159 | TGTAAAACGACGGCCAGT ATGACCTCAAACGGTGAGCA | CTTCCACATCCGAACCGACA | (GAGG)5 | 53 | 382-404 | |||
| 16 | BM-CiSSR-162 | TGTAAAACGACGGCCAGT GCTAGGGTTCCAGACTTCCAG | GATTTGGCCGATCGAAAGCC | (AAT)10(CAT)6 | 53 | 185-236 | |||
| 17 | BM-CiSSR-165 | TGTAAAACGACGGCCAGT AGCAACTTAAGGTCCTTCACGA | TTCTCTGCTCTGCTGTGCAT | (AAT)6 | 53 | 393-416 | |||
| 18 | BM-CiSSR-201 | TGTAAAACGACGGCCAGT CAACAGTACCTGATGGTCCG | TTCTGAAATCCAGTCCCCTC | (AAAAGA)6 | 55 | 259-270 | In this study | ||
| 19 | BM-CiSSR-202 | TGTAAAACGACGGCCAGT CCCTCTTCAAGAACTGAGCC | CACCAGCTGTTTGCTGTTTT | (AAAT)5 | 55 | 281-293 | |||
| 20 | BM-CiSSR-203 | TGTAAAACGACGGCCAGT ACAACGCACCAAGTCAATGT | GTTGCGTCATCCATTTTGTC | (AGCC)5 | 55 | 241-257 | |||
| 21 | BM-CiSSR-204 | TGTAAAACGACGGCCAGT CCATGACCCACTTTCCCTAC | ATTCGGGTAGGTTGAAATCG | (ACCCG)4 | 55 | 326-365 | |||
| 22 | BM-CiSSR-208 | TGTAAAACGACGGCCAGT GGATGCTTGGCCTGATTTAT | ATTGTCACCGAAGCACCATA | (TAA)7 | 55 | 340-352 | |||
| 23 | BM-CiSSR-210 | TGTAAAACGACGGCCAGT GCCAGGATTGAAGGGTTTTA | TGTGAACAAGGGCAACAGAT | (TTTAT)4 | 55 | 333-366 | |||
| 24 | BM-CiSSR-217 | TGTAAAACGACGGCCAGT ATAATGGAAGCGTCGGATTC | GCCTAACGGCCAGAGTTTAC | (CAG)10 | 55 | 313-327 | |||
| 25 | BM-CiSSR-218 | TGTAAAACGACGGCCAGT TATGTCTACTGGTCGCAGGC | GTTGTCCCCTTGATACCACC | (GGGAT)4 | 55 | 236-444 | |||
| 26 | BM-CiSSR-221 | TGTAAAACGACGGCCAGT GGTCCTTTGGAGAAGGTTGA | CATGACCAAATGTCGGGTTA | (TAAT)5 | 55 | 319-332 | |||
| 27 | BM-CiSSR-224 | TGTAAAACGACGGCCAGT AACCCCTTGTCAAGTGATCC | TCTTCTTCAGTTGGTGCCTG | (CAC)7 | 55 | 294-304 | |||
| 28 | BM-CiSSR-225 | TGTAAAACGACGGCCAGT GTAAGGGGTTGTGAGGCAAT | CAACAGGTTTCGACCATGAC | (GCT)8 | 55 | 322-353 | |||
| 29 | BM-CiSSR-226 | TGTAAAACGACGGCCAGT ATTAAGGCTGGAAATGCCAC | ATTCTGCTGACGCTTCAATG | (ATT)9 | 55 | 389-401 | In this study | ||
| 30 | BM-CiSSR-230 | TGTAAAACGACGGCCAGT TCCATCAGCCATTCCATCTA | ATCTGAACCCTCCAATCCTG | (TTC)9 | 55 | 276-302 | |||
| 31 | BM-CiSSR-238 | TGTAAAACGACGGCCAGT ACTATGCGGCTCGAACTTTT | TCACCTTCACAACCGAACAT | (CTT)11 | 55 | 269-277 | |||
| 32 | BM-CiSSR-239 | TGTAAAACGACGGCCAGT ACATGCCATAGGAAGCAACA | CACCTTCTCATCAATCACGG | (AAG)7 | 55 | 395-408 | |||
| 33 | BM-CiSSR-240 | TGTAAAACGACGGCCAGT GCTGCTGCTGCTAGTTTGTC | AAAGATGGCAATGGGTTAGG | (TCA)7 | 55 | 429-432 | |||
| 34 | BM-CiSSR-241 | TGTAAAACGACGGCCAGT GGTCCTTTGGAGAAGGTTGA | CATGACCAAATGTCGGGTTA | (TAAT)5 | 55 | 296-301 | |||
| 35 | BM-CiSSR-243 | TGTAAAACGACGGCCAGT CCATCCCTGTAAATTCCACC | ATTGGTCGTTTCCTTTCCTG | (AAAAT)4 | 55 | 319-323 | |||
| 36 | BM-CiSSR-245 | TGTAAAACGACGGCCAGT TTTCCCAGGAGCTTACCAAG | TGCGTTCCATGGTCAGTATT | (AAC)6 | 55 | 338-347 | |||
| 37 | BM-CiSSR-246 | TGTAAAACGACGGCCAGT CCCTAGGGAAATTTGGGAAT | GCACTCGAGAGTTCTCGTTAAG | (CAT)11 | 55 | 310-375 | |||
| 38 | BM-CiSSR-247 | TGTAAAACGACGGCCAGT ATCTGTGTTTGGTCGCATGT | GGAAGATTACCGGACTTGGA | (GAA)6 | 55 | 409-427 | |||
| 39 | BM-CiSSR-248 | TGTAAAACGACGGCCAGT GCTCGGTTCTTGCATACTGA | GTCTGCAAACCCTGTTGATG | (TGA)6 | 55 | 316-327 | |||
| 40 | BM-CiSSR-249 | TGTAAAACGACGGCCAGT ATGGGCAAGAACAGGAAATC | CCATAGGATTTGCATGAGGA | (GGA)6 | 55 | 321-331 | |||
| 41 | BM-CiSSR-253 | TGTAAAACGACGGCCAGT AATTTCCTGCTCCAAACCAG | TCCAACAACTTGAACACGGT | (TAA)14 | 55 | 319-355 | |||
| 42 | BM-CiSSR-254 | TGTAAAACGACGGCCAGT TAAAATCCCTCGGAAACAGG | CTTTGCATGTTCAACGTTCC | (ATC)6 | 55 | 234-313 | |||
| 43 | BM-CiSSR-256 | TGTAAAACGACGGCCAGT CTCTCTGAACCTGACACCGA | TTTTCTCCACCCTTTCAACC | (GAT)7 | 55 | 290-322 | |||
| 44 | BM-CiSSR-258 | TGTAAAACGACGGCCAGT GGTAAGCACCTGCAAACTGA | ATTATGCAATTCCTCCTGGC | (TATG)6 | 55 | 350-374 | |||
| 45 | BM-CiSSR-260 | TGTAAAACGACGGCCAGT TCATCTGAACGGACCACAAT | TAACTGCACTTGCTTCCCTG | (TTC)6 | 55 | 381-384 | |||
| 46 | BM-CiSSR-262 | TGTAAAACGACGGCCAGT CAGTTTCATCCCACTGATGC | ACCAAGCGTCCTTAACAACA | (ATT)6 | 55 | 181-205 | |||
| 47 | BM-CiSSR-264 | TGTAAAACGACGGCCAGT AGGGGTGCTGAGCATAAAAT | ATACCCCGTCGTGGAATTAG | (TAA)9 | 55 | 415-442 | |||
| 48 | BM-CiSSR-266 | TGTAAAACGACGGCCAGT CGTAGCCAAAACTCCCAAAT | CCGAAGATGGAGGGAACTAA | (GAA)8 | 55 | 293-342 | |||
| 49 | BM-CiSSR-268 | TGTAAAACGACGGCCAGT TCTGTGGCTCACTTCACTCC | GAAGACGACAGATGCTGGAA | (TCAC)7 | 55 | 355-365 | |||
| 50 | BM-CiSSR-270 | TGTAAAACGACGGCCAGT TGCTGTAAGTGCAGTGCAAA | GGGACGAGCATCTTCCTTTA | (TATG)5 | 55 | 342-347 | |||
| 51 | BM-CiSSR-271 | TGTAAAACGACGGCCAGT CCCCCAAAATGCTGAGTAGT | AAAGGGAGAGAGTTGGCTGA | (TATG)13 | 55 | 382-422 | |||
| 52 | BM-CiSSR-272 | TGTAAAACGACGGCCAGT ATAGGTCCCCACAATGGAAA | GGGCATAAATGAATTGGGTC | (GAA)8 | 55 | 405-419 | |||
| 53 | BM-CiSSR-273 | TGTAAAACGACGGCCAGT GCATCATACGTTCAAGCCAC | TCTTGTGCTCTCCTGTGACC | (CAT)12 | 55 | 298-336 | |||
yTa indicates annealing temperature.
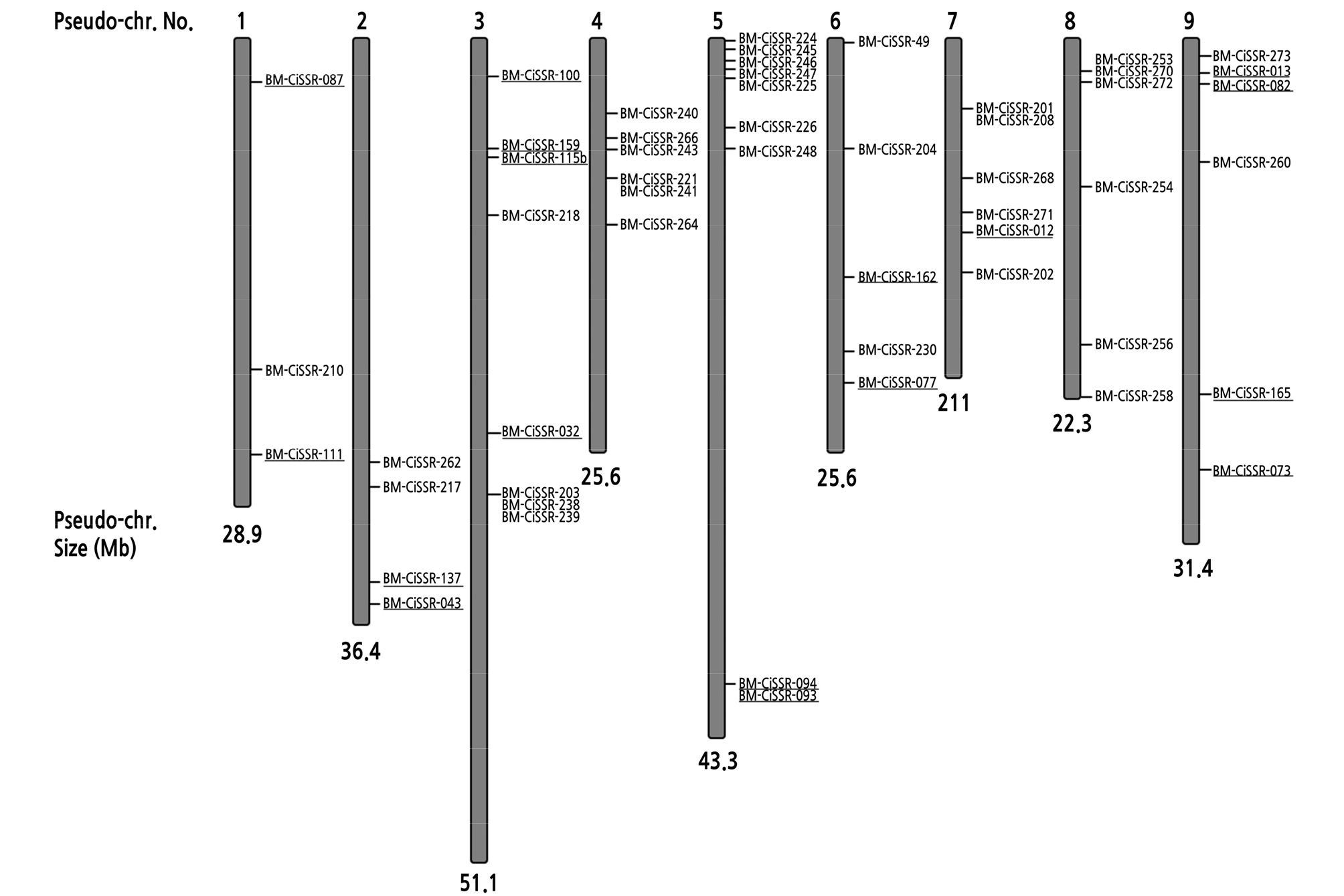
Fig. 1.
Physical map of 53 microsatellite loci on the nine pseudochromosomes of C. clementina cv. Clemenules. The underlined markers are 17 polymorphic microsatellites developed previously (Woo et al., 2019). Pseudo-chr. = pseudochromosome.
Identification of the Citrus Varieties Bred in Korea using Polymorphic Microsatellite Markers
Up to now, 34 citrus varieties were developed from bud sport mutants, nucellar mutants, and traditional genetic hybridizations by breeders at governmental institutes and breeding companies as well as private breeders in Korea (Table 2). Fifty-three microsatellite markers were applied to an available 32 Korean citrus varieties to determine their genotypes. Genetic characteristics of microsatellite loci based on the genotype analysis of 32 accessions are summarized in Table 3. A total of 245 alleles, ranging from 2 (BMCi-SSR084) to 11 (BMCi-SSR012) per locus, were observed among the 32 citrus accessions, with an average of five alleles per locus. Major allele frequency (MAF) varied from 0.22 (BMCi-SSR012) to 0.98 (BM-CiSSR-245). The average genetic diversity (GD) value was 0.53, ranging from 0.03 (BM-CiSSR-245) to 0.87 (BMCi-SSR012), and average polymorphism information content (PIC) value was 0.47, from 0.14 (BM-CiSSR-224) to 0.85 (BMCi-SSR012). The average observed heterozygosity (HO) was 0.47, with the lowest value in BM-CiSSR-245 and BM-CiSSR-270 (0.03) and the highest value in BM-CiSSR-262 (1.00).
Table 2.
Citrus varieties developed in Korea
yAgricultural Research and Extension Services, Jeju Special Self-Governing Province, Korea.
xHannong Bio Industry Corp., Korea.
wJenong Co., Ltd., Korea.
*Not available.
Table 3.
Characteristics of 53 microsatellite loci based on the genotype analysis of 32 citrus varieties developed in Korea
A total of 245 alleles derived from the 53 polymorphic microsatellite loci were used to evaluate genetic relationships among the 32 varieties. A UPGMA dendrogram was constructed based on the genetic similarity matrices among the accessions. Fig. 2 illustrates the results of the cluster analysis based on microsatellite data. The resulting dendrogram revealed that the accessions used in the analysis could be distinctly classified into lemon and non-lemon groups. The microsatellite markers clearly discriminated most of the citrus varieties developed in Korea, except for ‘Tamnaneunbong’ (TNB), ‘Saebyeolbong’ (SBB), and ‘Suneat’ (SNE), which originated from nucellar offspring (TNB and SBB) and bud sport mutation (SNE) of Citrus hybrid ‘Shiranuhi’ (Table 2). Nucellar embryos develop from somatic embryos derived from maternal nucellar tissue surrounding the sexual embryo sac. This process is a kind of apomixis and is widely found in many citrus varieties. The offspring derived from nucellar embryony in citrus possesses the same genetic constitution as the female parent (Kepiro and Roose, 2007; Zhang et al., 2018). Our results indicate that somatic mutants, including nucellar offspring, originating from an identical parental plant have identical or very similar genetic constitutions to each other.

In conclusion, we developed 53 polymorphic microsatellite markers covering nine pseudochromosomes of citrus and applied them to the identification of citrus varieties bred in Korea. The microsatellite markers clearly discriminated most of the citrus varieties, whereas somatic mutant varieties that were derived by nucellar embryony or bud sport mutation from a parental plant showed identical genotypes on all loci investigated due to identical or very similar genetic constitution. The microsatellite markers developed in this study will be a useful molecular tool to protect breeders’ rights and guarantee the quality of nursery plants in the citrus industry.
